ribóz-5-foszfátból és ATP-ből a PRPP-szintetáz (ribóz-foszfát pirofoszfokináz) által PRPP (5-foszforibozil-1-pirofoszfát) és AMP képződik
az újonan szintetizálódó purin gyűrű a PRPP által szolgáltatott ribóz-5-foszfáton fog felépülni
Purinok bioszintézise inozinátigPurinok bioszintézise IMP-től AMP és GMP képződéséig
PRPP-ből IMP (inozinát, inozin-monofoszfát) képződése: 11 vagy 10 lépésben
PRPP-ből és glutaminból a PRPP-glutamin-amidotranszferáz 5-foszfo--D-ribozilamint készít miközben glutamát és PPi lép ki a reakció során
5-foszfo--D-ribozilaminból, glicinből és ATP-ből a GAR-szintetáz (glicinamid ribonukleotid szintetáz) GAR-t készít (glicinamid ribonukleotid) miközben ADP és Pi lép ki
GAR-ból és N10-formil-H4-folátból (N-formil-tetrahidrofolát) a GAR-transzformiláz FGAR-t (formilglicinamid ribonukleotid) készít tetrahidrofolát kilépése közben
FGAR-ból, glutaminból és ATP-ből az FGAR-amidotranszferáz FGAM-ot (formilglicinamidin ribonukleotid) csinál miközben glutamát, ADP és Pi képződik
FGAM-ból és ATP-ből az FGAM-cikláz (AIR szintetáz) AIR-t (5-aminoimidazol ribonukleotid) készít víz, ADP és Pi kilépése közben
AIR-ból két módon keletkezhet CAIR:
Két egymást követő reakcióval:
AIR-ból, ATP-ből és HCO3--ból N5-CAIR-szintetáz C-t készít (N5-karboxiaminoimidazol ribonukleotid), ADP és Pi lép ki
N5-CAIR-ból az N5-CAIR-mutáz által CAIR (karboxiaminoimidazol ribonukleotid) képződik
Egy reakcióval:
AIR-ból CO2 hozzáadásával az AIR-karboxiláz CAIR-t készít
CAIR-ból IMP:
CAIR-ból, aszpartát és ATP hozzáadásával a SAICAR-szintetáz (N-szukcinil-5-aminoimidazol-4-karboxiamid ribonukleotid szintetáz) SAICAR-t képez ADP és Pi kilépésével
SAICAR-ból a SAICAR-liáz AICAR-t (5-aminoimidazol-4-karboxiamid ribonukleotid) képez fumarát kilépésével
AICAR-ból N10-formil H4-folát hozzáadásával az AICAR-transzformiláz FAICAR-t (N-formilaminoimidazol-4-karboxiamid ribonukleotid) készít tetrahidrofolát kilépése közben
FAICAR-ból IMP-szintáz IMP-t (inozinát) készít vízkilépéses reakció során
IMP-ből képződhet AMP (adenilát, adenozin-monofoszfát) és GMP (guanilát, guanozin-monofoszfát):
AMP képződése:
IMP-ből, aszpartát és GTP hozzáadásával az adeniloszukcinát-szintetáz adeniloszukcinátot készít miközben GTP és Pi lép ki
adeniloszukcinátból az adeniloszukcinát-liáz AMP-t képez fumarát kilépése közben
GMP képződése:
IMP-ből, víz és NAD+ hozzáadásával az IMP-dehidrogenáz XMP-t (xantilát) készít miközben NADH+H+ lép ki
XMP-ből, glicin, víz és ATP hozzáadásával a XMP-glutamin-amidotranszferáz GMP-t képez glutamát, AMP és PPi kilépése közben
Példa nukleozid-difoszfát és -trifoszfát keletkezéseére
Nukleozid-difoszfátok és trifoszfátok keletkezése:
AMP-ből és GMP-ből nukleozid-monofoszfát-kinázok és nukleozid-difiszfát-kinázok képezik őket
ATP a magas energiájú foszfátcsoportforrás ezekben a reakciókban
Adenin-foszforibozil-transzferáz és hipoxantin-guanin-foszforibozil-transzferáz reakciók
Bizonyos sejtek (pl. eritrociták) nem tudnak de novo purin nukleotidokat szintetizálni és felvenni sem. Viszont purin nukleozidokat exogén fel tudnak venni, amikből ezen útvonal segítségével látják el a sejtek nukleotid szükségletét.
PRPP + purin bázis
PRPP-ből és adeninből az adenin-foszforibozil-transzferáz AMP-t hoz létre PPi kilépéssel
PRPP-ből és hipoxantinból vagy guaninból a hipoxantin-guanin-foszforibozil-transzferáz IMP-t vagy GMP-t hoz létre PPi kilépésével
purin-ribonukleozid ATP-vel foszforilálódik és nukleozid-monofoszfátok keletkeznek
adenozinból az adenozin-kináz csinál AMP-t (ATP ADP)
guanozinból a guanozin-kináz csinál GMP-t (ATP ADP)
Fájl:9 karbamoil-foszfát szintetáz.JPGKarbamoil-foszfát szintetáz II. citoszólikus enzim által katalizált reakciókaszpartátból és karbamoil-foszfátból az aszpartát-transzkarbamiláz készít N-karbamoil aszpartátot, miközben Pi lép ki
glutaminból, HCO3--ből és két ATP-ből a karbamoil-foszfát szintetáz II. (citoszólban lévő enzim) szintetizálja a karbamoil-foszfátot:
A purin és pirimidin nukleotidok lebontása a foszfátcsoportok eltávolításával (víz belépés mellett) kezdődik nukleotidázok által. Ennek eredményeként adenozin, guanozin, inozin, citidin és uridin keletkezik.
A dezoxiribonukleotidok kialakulásáért a ribonukleotid-reduktáz a felelős. Ez csak olyan sejtekben aktív, melyek osztódni akarnak. A ribóz második szénatomján lévő hidroxilcsoport redukálódik és oxigénből víz keletkezik. Az enzimrendszer több komponensből épül fel és SH csoportokat is tartalmaz, amik az enzim működése közben oxidálódnak. Kofaktora a tioredoxin. Ennek visszaredukálását eukariótákban a glutation-reduktáz végzi, amelyhez NADPH-t használ hidrogéndonorként. Az enzim az összes nukleotidot szubsztrátnak tekinti, ha azok nukleozid-difoszfát (ADP, CDP, UDP, GDP) formában vannak jelen.
Súlyos kombinált immunhiány: autoszomális recesszív
ok: adenozin-deamináz hiány
nincs adenozin inozin átalakulás
nincs dAMP dIMP átalakulás
az enzim hiánya az adenozin, dAMP felhalmozódásához vezet
dAMP átalakul dADP-vé és dATP-vé, amelyek felhalmozódnak a sejtekben
dATP gátolja a ribonukleotid reduktázt, vagyis csökkenti a ribonukleotidok dezoxiribonukleotiddá alakulását, ami csökkenti a DNS szintézist a B és T sejtekben
Fájl:2 ribonukleotid reduktáz 1.JPGRibonukleotid-reduktáz enzim működésének mechanizmusaB és T sejtek hiánya a humorális és sejtes immunitás csökkenése miatt visszatérő bakteriális, vírus, gomba és protozoon fertőzésekhez vezet
A gyorsan növekvő daganatos sejtek és baktériumok DNS és RNS szintézise magas nukleotid szintet igényel, ezért a gyorsan növekvő daganatos sejtek és baktériumok érzékenyebbek a nukleotid bioszintézis gátlókra. Azonban nem csak a nem kedvező, hanem a fontos és hasznos funkciókat szolgáló sejtek is károsodnak, mivel ugyan a leggyorsabban osztódó sejtek károsodnak a legnagyobb mértékben, de nem kizárólag. Ezért pl. vérképző-, bélhám- és ivarsejteket és immunrendszert is érinti az ilyen jellegű kezelés.
Alapja, hogy a természetes anyagcseretermékekhez hasonló szerkezetű vegyületeket használnak, amik így gátolhatják az adott enzimet.
Folát analógok (pl. aminopterin, metotrexát, trimetoprim)
kompetitíven gátolja a dihidrofolát reduktázt, ami a dihidrofolát tetrahidrofáláttá való redukcióját katalizálja
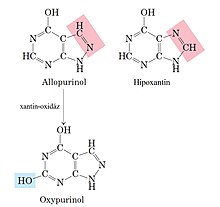
Fájl:1 köszvény.jpgKöszvénytetrahidrofolát kell a timidin és purin nukleotidok szintéziséhez
alkalmazása:
Mola-betegség
leukémiák
limfómák
oszteogén szarkóma
reumatoid artritiszKöszvény kezelése
5-fluorouracil (5-FU)
tumorsejtek 5-fluorodeoxiuridin-monofoszfáttá (FdUMP) alakítják, ami irreverzibilisen gátolja a timidilát-szintázt, és akadályozza a deoxitimidin-monofoszát (dTMP) szintézisét deoxiuridin-monofoszfátból (dUMP)
alkalmazása:
emlő-, gyomor- és vastagbélrák
Hidroxiurea
gátolja a ribonukleotid-reduktázt, ami a ribonukleotidokat dezoxiribonukleotiddá alakítja át
dezoxiribonukleotidok mennyiségének csökkenésével csökken a DNS szintézis is
alkalmazása:
krónikus mielogén leukémiaFolát analógok és fluorouracil alkalmazása a daganatterápiában
sarlósejtes anaémia
6-merkaptopurin és származéka (pl. Imuran)
a hipoxantin szerkezeti analógja
foszforibozilálása hipoxantin-guanin-foszforibozil-transzferáz jelenlétében tioinozinsavat eredményez, mely az inozinsav adeniláttá és guaniláttá történő átalakulását gátolja
Glutamin analógok
Citozin-arabinozid
citozintól annyiban tér el, hogy ribóz helyett arabinóz a cukorkomponense
a sejtekben foszforilálódik és így, mint a dCTP analógja, gátolja a DNS-szintézist
Lehninger Principles of Biochemistry: David L. Nelson, Michael M. Cox: Lehninger Principles of Biochemistry. New York, W.H. Freeman and Company, 2008. 5thISBN 978-0-7167-7108-1
Biokémia - Gyors segítség a sikeres vizsgához: Edward F. Goljan (szerk.): Biokémia - Gyors segítség a sikeres vizsgához. Budapest, Medicina Könyvkiadó Zrt., 2009. ISBN 978-963-226-254-3
Orvosi biokémia: Ádám Veronika (szerk.): Orvosi biokémia. Budapest, Medicina Könyvkiadó Zrt., 2006. ISBN 963-242-902-8
Szerves és bioorganikus kémia: Gergely Pál, Penke Botond, Tóth Gyula: Szerves és bioorganikus kémia. Budapest, Alliter Kiadói és Oktatásfejlesztő Alapítvány, 2006. ISBN 963-87040-4-7

























{kind=link}
{kind=link}
{kind=link}
{kind=link}
